Insects
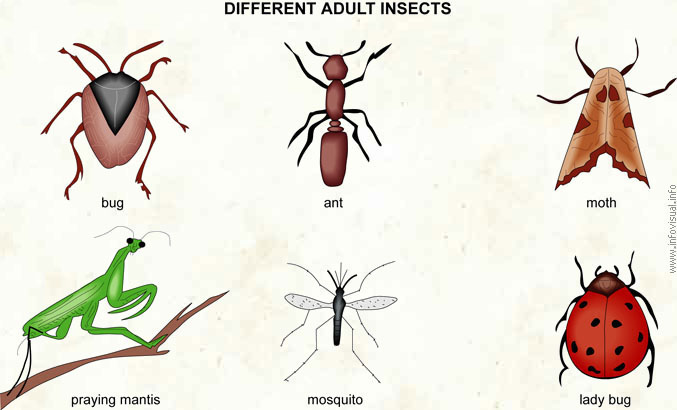
Bug: heteropterous insect that releases a sickening odor.
Ant: wingless social insect.
Moth: butterfly-like nocturnal insect.
Praying mantis: carnivorous insect.
Mosquito: insect of which the female bites humans or animals to feed off their blood.
Lady bug: aphid-eating coleopteron, also called ladybugs.
Photo :

Insects, Class Insecta are a major group of arthropods and the most diverse group of animals on the Earth, with over a million described species, more than half of all known living organisms with estimates of undescribed species as high as 30 million, thus potentially representing over 90% of the differing life forms on the planet. Insects may be found in nearly all environments on the planet, although only a small number of species occur in the oceans, a habitat dominated by another arthropod group, the crustaceans.
There are approximately 2,000 praying mantis, 5,000 dragonfly species, 20,000 grasshopper, 82,000 true bug, 120,000 fly, 110,000 bee, wasp and ant, 170,000 butterfly and moth, and 360,000 beetle species described to date. Estimates of the total number of current species, including those not yet known to science, range from two million to fifty million, with newer studies favouring a lower figure of about six to ten million. Adult modern insects range in size from a 0.139 mm (0.00547 in) fairyfly (Dicopomorpha echmepterygis) to a 55.5 cm (21.9 in) long stick insect (Phobaeticus serratipes). The heaviest documented insect was a Giant Weta of 70 g (2½ oz), but other possible candidates include the Goliath beetles Goliathus goliatus, Goliathus regius and Cerambycid beetles such as Titanus giganteus, though no one is certain which is truly the heaviest.
Insects possess segmented bodies supported by an exoskeleton, a hard outer covering made mostly of chitin. The segments of the body are organized into three distinctive but interconnected units, or tagmata; a head, a thorax, and an abdomen. The head supports a pair of sensory antennae, a pair of compound eyes, one to three simple eyes ("ocelli") and three sets of variously modified appendages that form the mouthparts. The thorax has six segmented legs (one pair each for the prothorax, mesothorax and the metathorax segments making up the thorax) and two or four wings (if present in the species). The abdomen (made up of eleven segments some of which may be reduced or fused) has most of the digestive, respiratory, excretory and reproductive internal structures.

Their nervous system can be divided into a brain and a ventral nerve cord. The head capsule, made up of six fused segments, has six pairs of ganglia. The first three pairs are fused into the brain, while the three following pairs are fused into a structure called the subesophageal ganglion. The thoracic segments have one ganglion on each side, which are connected into a pair, one pair per segment. This arrangement is also seen in the abdomen but only in the first eight segments. Many species of insects have reduced numbers of ganglia due to fusion or reduction. Some cockroaches have just six ganglia in the abdomen, whereas the wasp Vespa crabro has only two in the thorax and three in the abdomen. And some, like the house fly Musca domestica, have all the body ganglia fused into a single large thoracic ganglion.
Until very recently, no one had ever documented the presence of nociceptors (the cells that detect and transmit sensations of pain) in insects, though recent findings of nociception in larval fruit flies challenges this and raises the possibility that some insects may be capable of feeling pain.
Insect respiration is accomplished without lungs, using a system of internal tubes and sacs through which gases either diffuse or are actively pumped, delivering oxygen directly to the adjoining body tissues (see Invertebrate trachea). Since oxygen is delivered directly, the circulatory system is not used to carry oxygen, and is therefore greatly reduced; it has no closed vessels (i.e., no veins or arteries), consisting of little more than a single, perforated dorsal tube which pulses peristaltically, and in doing so helps circulate the hemolymph inside the body cavity. Air is taken in through spiracles, openings on the sides of the abdomen.
Most higher insects have two pairs of wings located on the second and third thoracic segments. Insects are the only invertebrates to have developed flight, and this has played an important part in their success. The winged insects, and their wingless relatives, make up the subclass Pterygota. Insect flight is not very well understood, relying heavily on turbulent aerodynamic effects. The primitive insect groups use muscles that act directly on the wing structure. The more advanced groups making up the Neoptera have foldable wings and their muscles act on the thorax wall and power the wings indirectly. These muscles are able to contract multiple times for each single nerve impulse, allowing the wings to beat faster than would ordinarily be possible.
Their outer skeleton, the cuticle, is made up of two layers; the epicuticle which is a thin and waxy water resistant outer layer and contains no chitin, and another layer under it called the procuticle. This is chitinous and much thicker than the epicuticle and has two layers, the outer being the exocuticle while the inner is the endocuticle. The tough and flexible endocuticle is built from numerous layers of fibrous chitin and proteins, criss-crossing each others in a sandwich pattern, while the exocuticle is rigid and sclerotized. The exocuticle is greatly reduced in many soft-bodied insects, especially the larval stages (e.g., caterpillars).
Most insects hatch from eggs, but some are ovoviviparous or viviparous, and all undergo a series of moults as they develop and grow in size. This manner of growth is necessitated by the inelastic exoskeleton. Moulting is a process by which the individual escapes the confines of the exoskeleton in order to increase in size, then grows a new and larger outer covering. In some insects, the young are called nymphs and are similar in form to the adult except that the wings are not developed until the adult stage. This is called incomplete metamorphosis and insects showing this are termed hemimetabolous. Holometabolous insects show complete metamorphosis, which distinguishes the Endopterygota and includes many of the most successful insect groups. In these species, an egg hatches to produce a larva, which is generally worm-like in form, and can be divided into five different forms; eruciform (caterpillar-like), scarabaeiform (grublike), campodeiform (elongated, flattened, and active), elateriform (wireworm-like) and vermiform (maggot-like). The larva grows and eventually becomes a pupa, a stage marked by reduced movement and often sealed within a cocoon. There are three types of pupae; obtect (the pupa is compact with the legs and other appendages enclosed), exarate (where the pupa has the legs and other appendages free and extended) and coarctate (where the pupa develops inside the larval skin). In the pupal stage, the insect undergoes considerable change in form to emerge as an adult, or imago. Butterflies are an example of an insect that undergoes complete metamorphosis. Some insects have even evolved hypermetamorphosis.

Many insects possess very sensitive and/or specialized organs of perception. Some insects such as bees can perceive ultraviolet wavelengths, or detect polarized light, while the antennae of male moths can detect the pheromones of female moths over distances of many kilometres. There is a pronounced tendency for there to be a trade-off between visual acuity and chemical or tactile acuity, such that most insects with well-developed eyes have reduced or simple antennae, and vice-versa. There are a variety of different mechanisms by which insects perceive sound, and it is by no means universal; the general pattern, however, is that if an insect can produce sound, then it can also hear sound, though the range of frequencies they can hear is often quite narrow (and may in fact be limited to only the frequency that they themselves produce). Some nocturnal moths can perceive the ultrasonic emissions of bats, a mechanism which helps them avoid predation. Certain predatory and parasitic insects can detect the characteristic sounds made by their prey/hosts. Bloodsucking insects have special sensory structures that can detect infrared emissions, and use them to home in on their hosts.
A few such insects also have a well-developed number sense, among the solitary wasps that provision with a single species of prey. The mother wasp lays her eggs in individual cells and provides each egg with a number of live caterpillars on which the young feed when hatched. Some species of wasp always provide five, others twelve, and others as high as twenty-four caterpillars per cell. The number of caterpillars is different among species, but it is always the same for each sex of larvae. The male solitary wasp in the genus Eumenes is smaller than the female, so the mother of one species supplies him with only five caterpillars; the larger female receives ten caterpillars in her cell. She can in other words distinguish between both the numbers five and ten in the caterpillars she is providing and which cell contains a male or a female.
A few insects, such as members of the families Poduridae and Onychiuridae (Collembola), Mycetophilidae (Diptera), and the beetle families Lampyridae, Phengodidae, Elateridae and Staphylinidae are bioluminescent. The most familiar group are the fireflies, beetles of the family Lampyridae. Some species are able to control this light generation to produce flashes. The function varies with some species using them to attract mates, while others use them to lure prey. Cave dwelling larvae of Arachnocampa (Mycetophilidae, Fungus gnats) glow to lure small flying insects into sticky strands of silk. Some fireflies of the genus Photuris mimic the flashing of female Photinus species to attract males of that species, which are then captured and devoured. The colours of emitted light vary from dull blue (Orfelia fultoni, Mycetophilidae) to the familiar greens and the rare reds (Phrixothrix tiemanni, Phengodidae).
Most insects except some species of cave dwelling crickets are able to perceive light and dark. Many species have acute vision capable of detecting minute movements. The eyes include simple eyes or ocelli as well as compound eyes of varying sizes. Many species are able to detect light in the infrared, ultraviolet as well as the visible light wavelengths. Colour vision has been demonstrated in many species and phylogenetic analysis suggests that the basic bauplan of UV-green-blue trichromacy existed from at least the Devonian period.
Insects were the earliest organisms to produce sounds and to sense them. Soundmaking in insects is achieved mostly by mechanical action of appendages. In the grasshoppers and crickets this is achieved by stridulation. The cicadas have the loudest sounds among the insects and have special modifications to their body and musculature to produce and amplify sounds. Some species such as the African cicada, Brevisana brevis have been measured at 106.7 decibels at a distance of 50 cm (20 in). Some insects, such as the hawk moths and Hedylid butterflies, can hear ultrasound and take evasive action when they sense detection by bats. Some moths produce ultrasound clicks and these were earlier thought to have a role in jamming the bat echolocation, but it was subsequently found that these are produced mostly by unpalatable moths to warn bats, just as warning colourations are used against predators that hunt by sight. These calls are also made by other moths involved in mimicry. Very low sounds are also produced in various species of Neuroptera, Lepidoptera (butterflies and moths), Coleoptera and Hymenoptera produced by the mechanical actions of movement often aided by special microscopic stridulatory structures.

Some species use vibrations for communicating within members of the same species, such as to attract mates as in the songs of the shield bug Nezara viridula while it can also be used to communicate between entirely different species, such as between ants and myrmecophilous lycaenid caterpillars. In addition to the use of sound for communication, a wide range of insects have evolved chemical means for communication. These chemicals, termed semiochemicals, are often derived from plant metabolites include those meant to attract, repel and provide other kinds of information. While some chemicals are targeted at individuals of the same species, others are used for communication across species. The use of scents is especially well known to have developed in social insects.
Insects are the only group of invertebrates to have developed flight. The evolution of insect wings has been a subject of debate. Some proponents suggest that the wings are para-notal in origin while others have suggested they are modified gills. In the Carboniferous age, some of the Meganeura dragonflies had as much as a 50 cm (20 in) wide wingspan. The appearance of gigantic insects has been found to be consistent with high atmospheric oxygen. The percentage of oxygen in the atmosphere found from ice core-samples was as high as 35% compared to the current 21%. The respiratory system of insects constrains their size, however the high oxygen in the atmosphere allowed larger sizes. The largest flying insects today are much smaller and include several moth species such as the Atlas moth and the White Witch (Thysania agrippina).
Insect flight has been a topic of great interest in aerodynamics due partly to the inability of steady-state theories to explain the lift generated by the tiny wings of insects. In addition to powered flight, many of the smaller insects are also dispersed by winds. These include the aphids which are often transported long distances by low-level jet streams.
Many adult insects use six legs for walking and have adopted a tripedal gait. The tripedal gait allows for rapid walking whilst always having a stable stance and has been studied extensively in cockroaches. The legs are used in alternate triangles touching the ground. For the first step the middle right leg and the front and rear left legs are in contact with the ground and move the insect forward, whilst the front and rear right leg and the middle left leg are lifted and moved forward to a new position. When they touch the ground to form a new stable triangle the other legs can be lifted and brought forward in turn and so on. The purest form of the tripedal gait is seen in insects moving at speed. However, this type of locomotion is not rigid and insects can adapt a variety of gaits; for example, when moving slowly, turning, or avoiding obstacles, four or more feet may be touching the ground. Insects can also adapt their gait to cope with the loss of one or more limbs.
Cockroaches are amongst the fastest insect runners and at full speed actually adopt a bipedal run to reach a high velocity in proportion to their body size. As Cockroaches move extremely rapidly, they need recording at several hundred frames per second to reveal their gait. More sedate locomotion is also studied by scientists in stick insects Phasmatodea. A few insects have evolved to walk on the surface of the water, especially the bugs of the family, Gerridae, also known as water striders. A few species of ocean-skaters in the genus Halobates even live on the surface of open oceans, a habitat that has few insect species.
Animation : World insects
Thanks to YouTube for allowing us to watch this video.